
抗体(Antibody,Ab)是介导体液免疫的重要效应分子,是免疫系统在外来颗粒或生物体的抗原刺激下,由B细胞或记忆B细胞增殖分化成的浆细胞所产生的、可与相应的抗原发生特异性结合的免疫球蛋白(Immunoglobulin,Ig),主要分布在血清中,也分布于组织液、外分泌液及某些细胞膜表面。Ab的基本结构是由两条完全相同的重链和两条完全相同的轻链通过二硫键连接呈的Y形单体,根据重链恒定区的差异可分为IgM、IgG、IgA、IgD和IgE五类同种型。
IgA与IgG简介
IgA是呼吸道、胃肠道等黏膜表面分泌物中的主要抗体类别,是机体抗原物质的主要来源。IgA具有抑制细菌、病毒与上皮细胞的黏附,中和细胞内外病毒和细菌毒素的功能,被认为是抵御病原体入侵的第一道防线。IgA有血清型和分泌型两种,血清IgA直接分泌到血液中,以单体形式存在,其中约90%是在骨髓中产生的IgA1单体。血清IgA分子量为160kD左右,占总Ig的10%–15%,浓度约为1.5~2.6mg/mL,仅次于IgG(9.5~12.5mg/mL),产生速率与IgG相似,但代谢速度比IgG快五倍。
分泌型IgA(SIgA)是外分泌液中的主要抗体类型,参与黏膜局部免疫,在局部抗感染中发挥重要作用,是机体抗感染的“边防军”。分泌型IgA(SIgA)主要以二聚体(dIgA)为主,在肠道、呼吸道、乳腺、唾液腺和泪腺等部位合成,经黏膜上皮细胞分泌至胃肠道和支气管分泌液、初乳、唾液和泪液中。SIgA通过特异性结合,阻止病原体黏附到细胞表面。
免疫球蛋白G(Immunoglobulin G, IgG)
IgG是二次免疫应答产生的主要同种型抗体,亲和力高、分布广泛,是机体抗感染的“主力军”。IgG在人出生后3个月开始合成,是血清和胞外液中含量最高的免疫球蛋白,约占血清总Ig的75%~80%,半衰期为20~23天,是半衰期最长的同种型Ig。基于重链恒定区,特别是CH1和CH3的结构、抗原和功能差异,IgG分为IgG1、IgG2、IgG3、IgG4四个亚类。二次免疫反应中,抗原诱导产生IgG亚型因抗原类型而异,蛋白质抗原通常诱导产生IgG1和IgG3抗体,多糖类抗原通常诱导产生IgG2和IgG4抗体。
IgA、IgG等同种型抗体是通过IgM同种型类别转换产生的。B细胞在Ig重链V区基因重排后其子代细胞中的重链V区基因保持不变,但位于VDJ基因座下游的是九个功能性CH基因则会发生重排,即同种型类别转换。Cμ、Cδ、Cγ3、Cγ1、Cα1、Cγ2、Cγ4、Cε、Cα2九个CH基因,除了Cδ外,每个CH基因的前面都有一个不能翻译的外显子和一个开关区(S区)。在同种型转换期间,中心细胞的CH区经历一系列DNA切割或重新连接事件,一旦接收到转换信号,DNA很可能被环出,中间未使用的CH外显子将被删除,选定的CH外显子与重排的VDJ重链外显子连接。VDJ重链外显子与新选定的外显子剪接产生mRNA,该mRNA被翻译成新的重链,导致B细胞从IgM转换为新的Ig类型。
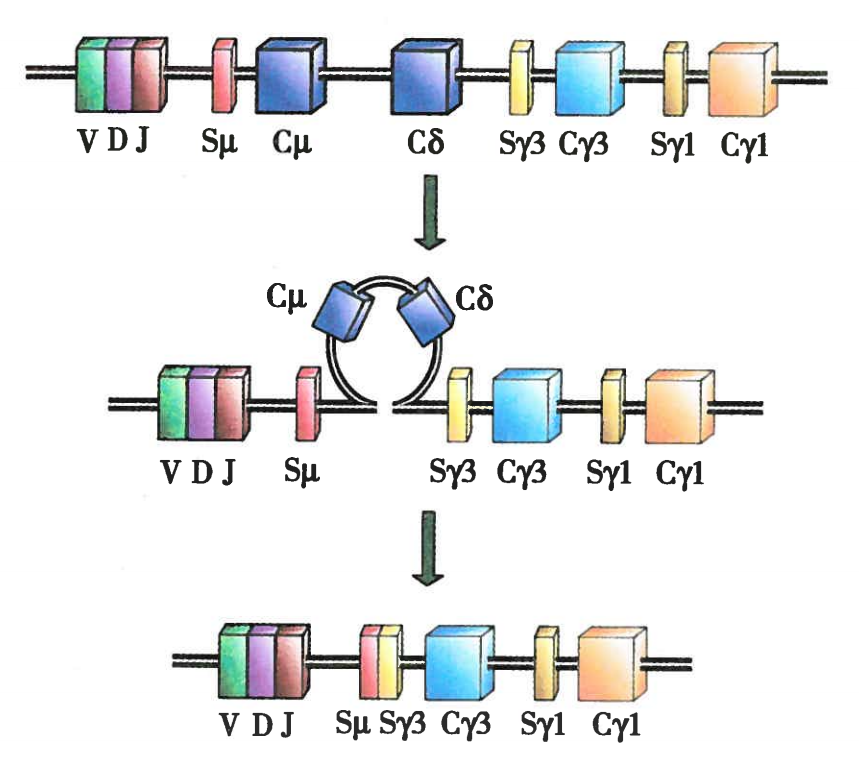
图1 Ig类别转换
注:Sμ、Sγ3、Sγ1分别为Cμ、Cγ3、Cγ1基因上游的转换区,在Ig类别转换为IgG3时,Sμ和、Sγ3重组,期间的序列包括Cμ、Cδ都被环出,从而使Ig类别转换为IgG3。
IgA和IgG之间的主要区别在于分泌多肽,IgA具有促进通过黏膜表面分泌的分泌多肽,而IgG没有。IgA具有J链(Joining chain)和分泌成分(Secretory component,SC)两种抗体辅助成分。J链是一种富含半胱氨酸的酸性糖蛋白,包含124个氨基酸,分子量约15kD,由浆细胞合成。J链通过尾部的倒数第二个Cys残基与单体IgA形成二硫键,聚合IgA。J链是IgA上皮细胞间的转运及其在黏膜表面分泌的先决条件,对IgA分泌至关重要。SC是多聚免疫球蛋白受体(pIgR)的胞外段,是一种由黏膜上皮细胞合成、分泌的含糖肽链,分子量约为75kD。多聚体IgA通过J链与pIgR结合,辅助SIgA由黏膜固有层,经黏膜上皮细胞转运,分泌到黏膜表面,最终pIgR蛋白水解并释放SC-pIgA复合物(SIgA),SIgA中的SC可保护其铰链区免受蛋白水解酶降解作用。
此外,单体IgA重链C端存在一个18个氨基酸组成的尾部,IgG重链中并没有这种结构。IgG、IgA抗体结合抗原表位的个数(即抗原结合价)也存在差异,单体IgG、IgA为2价,分泌型IgA(主要是二聚体IgA)为4价。
分泌型IgA可被转运到呼吸道和消化道黏膜表面,在黏膜局部免疫中发挥重要作用。黏膜上皮固有层中的局部浆细胞(PC)产生IgA。聚合IgA(主要是二聚体IgA)通过J链与聚合免疫球蛋白受体(pIgR)在基底外侧表面结合,形成的pIgR-IgA复合物被内吞并通过一系列囊泡穿过细胞被传递到顶端表面。在顶端表面的pIgR蛋白水解,释放SC-pIgA复合物(SIgA)或游离分泌成分(SC)。
IgG穿过胎盘的作用是一种重要的自然被动免疫机制。胎盘母体一侧的滋养层细胞表达一种IgG输送蛋白,称为新生Fc段受体(FcRn)。FcRn通过CH2-CH3结构域与IgG结合,介导母体IgG对新生儿的转胞吞作用,调节血清IgG水平。IgG与FcRn的结合严格依赖于pH,在体内酸性环境下与FcRn结合,通过胞吞作用转移到滋养层细胞内,并通过pH变化释放到进入胎儿血液循环中。FcRn结合可保护其免受溶酶体的破坏。
补体激活
补体级联激活是清除调理病原体的重要手段,抗体与相应抗原结合后,因构型改变而使其CH2和CH3结构域内的补体结合位点暴露,从而通过经典途径激活补体系统,产生多种补体的效应功能。IgG不同亚型补体激活能力不同,IgG4是唯一不能激活补体的IgG亚类,而IgG1、IgG2、IgG3对补体激活成分亲和力差异显著。IgG1和IgG3激活补体的能力较强,IgG2较弱,以C1q补体成分为例,IgG的亲和力大小依次为IgG3>IgG1>IgG2。相比于IgG,IgA是一种较弱的补体激活剂,可通过旁路途径激活补体系统。在抗原有限的情况下,通过阻断强补体激活剂IgG或IgM抗体的结合抑制补体激活。
婴幼儿被动免疫
人类初乳中分泌的IgA浓度非常高,含有针对多种微生物抗原的SIgA,婴儿可从母亲初乳中获得SIgA,在肠道中SIgA发挥分子“油漆”作用,干扰细菌运动并包围具有亲水外壳的病原体,阻止病原体进入肠上皮。SIgA通过抗体介导的表面抗原交联会凝集细菌,中和酶、毒素等细菌产物,再通过肠蠕动去除细菌聚集体。SIgA与黏液层形成抵抗病原微生物的天然屏障,共同防止病原微生物在黏膜上皮定植和渗透,避免感染。研究显示,母乳喂养可显著降低婴儿死于胃肠道和呼吸道感染的几率。
与IgA分子“油漆”作用不同,IgG可以穿过胎盘屏障,进入胎儿血液循环发挥被动免疫作用。IgG是唯一能通过胎盘的免疫球蛋白,对新生儿抗感染免疫至关重要。
表1 IgA、IgG主要理化性质及生物学功能
| 参数 | IgA | IgG |
|---|---|---|
| 开始合成时间 | 出生后4~6个月 | 出生后3个月 |
| 主要存在形式 | 单体/二聚体 | 单体 |
| 合成率[mg/(kg•d)] | 65 | 33 |
| 占总血清Ig比例 | 10~15% | 75~85% |
| 血清含量mg/mL | 1.5~2.6 | 9.5~12.5 |
| 半衰期(天) | 6 | 23 |
| 单体分子量(kD) | 160 | 150 |
| 重链 | α | γ |
| 亚类 | IgA1、IgA2 | IgG1、IgG2、IgG3、IgG4 |
| 抗原结合价 | 2,4 | 2 |
| 糖基化修饰 | 7% | 3% |
| 胎盘转运 | – | + |
| 结合SPA | – | + |
| 经典途径补体激活 | – | + |
| 旁路途径补体激活 | IgA1+ | IgG4+ |
| 主要作用 | 黏膜免疫 | 再次免疫应答 |
参考文献
[1]Schroeder HW Jr, Cavacini L. Structure and function of immunoglobulins[J]. J Allergy Clin Immunol., 2010,125(2 Suppl 2):S41-52.
[2]Macpherson, A., McCoy, K., Johansen, FE., et al. The immune geography of IgA induction and function[J].Mucosal Immunol, 2008: 11–22.
[3]医学免疫学[M].






南京德泰生物工程有限公司 Nanjing Detai Bioengineering Co.,Ltd. ©2024 All Rights Reserved
 苏公网安备32011202001300
苏公网安备32011202001300