
自然界中抗原结构复杂、种类繁多,并且含有多种表位。面对抗原的刺激,机体可以产生抗原表位特异性抗体,以及针对同一表位的不同类型抗体。
Ig基因重排
人的Ig重链基因由编码可变区的V基因片段(VH)、D基因片段(DH)、J基因片段(JH)以及编码恒定区的C基因片段组成,VH、DH和JH的基因片段数分别为45、23和6个。Κ或λ轻链基因可变区由V、J基因片段组成,Vκ和Jκ基因片段数分别为40和5个,Vλ和Jλ基因片段数分别为30和4个;重组C基因片段有9个,按照5’-Cμ-Cδ-Cγ3-Cγ1-Cα1-Cγ2-Cγ4-Cε-Cα2-3’的顺序排列。Cκ基因片段数只有1个,Cλ基因片段数有4个,按照5’-Cγ1-Cγ2-Cγ3-Cγ7-3’的顺序排列。Ig胚系基因以被分隔开的基因片段成簇存在,通过重组酶的作用,从众多V(D)J基因片段中各选择一个V片段、D片段(轻链无)、J片段,形成V-D-J(重链)或V-J(轻链)连接,再与C基因片段连接,编码形成完整的Ig多肽链,进一步加工、组装成有功能的BCR。在B细胞分化发育过程中,BCR基因片段发生重新排列和组合,产生数量巨大、能识别特异性抗原的BCR。
人类Ig基因片段可以产生多种V区基因片段组合,理论上仅重链V区的排列就可达45(VH)×23(VD)×6(VJ)=6210之多,轻重链组合多样性可达1.9×106。因V区基因片段使用频率及轻重链配对成功概率不同,实际组合数量要比理论上少一些,但仍具有丰富的多样性。此外,Ig基因片段还可以通过密码子错位、框架移位以及N序列插入等形式产生新的序列,从而增加BCR多样性。
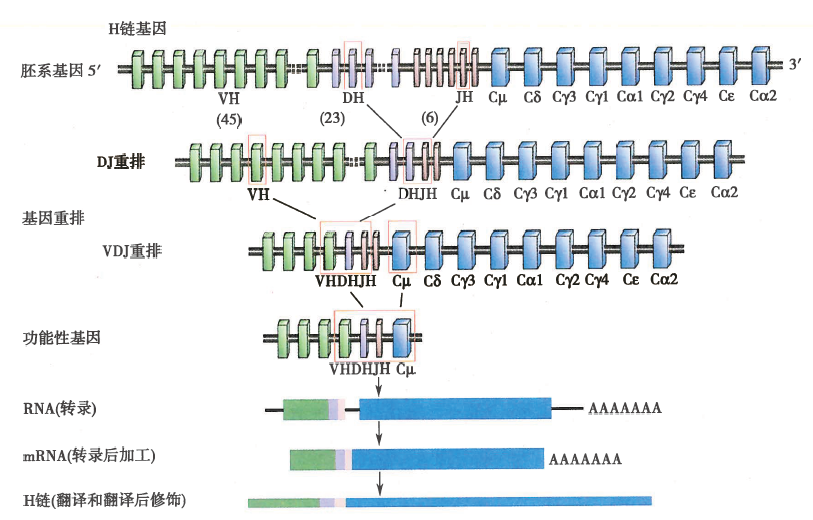
图1 免疫球蛋白重链基因重排及表达示意图
注:重链胚系基因经过重排先形成D-J连接,然后形成V-DJ连接,编码功能性V区基因。
抗体类别转换
2020年Cell Reports上发表了一篇题为“Antibody Isotype Switching as a Mechanism to Counter HIV Neutralization Escape”的文章。研究者发现在HIV感染的不同时段,体内的抗体同种型差异明显,同种型抗体是通过抗体类别转换重组(CSR)产生的,CSR是一种B细胞特异性DNA重排反应,且不同同种型的抗体对病毒具有不同的中和作用。其中IgG3和IgA1中和活性较好,IgG3类抗体能够通过抗体依赖的细胞毒作用(ADCC)发挥抗病毒作用。为了产生最佳的免疫反应,成熟B淋巴细胞通过将表达Ig重链恒定区(CH)的基因从Cμ交换到另一个下游CH基因,将IgM抗体转换为具有相同抗原特异性、不同效应功能的二级同种型(IgG、IgA 或IgE)抗体。
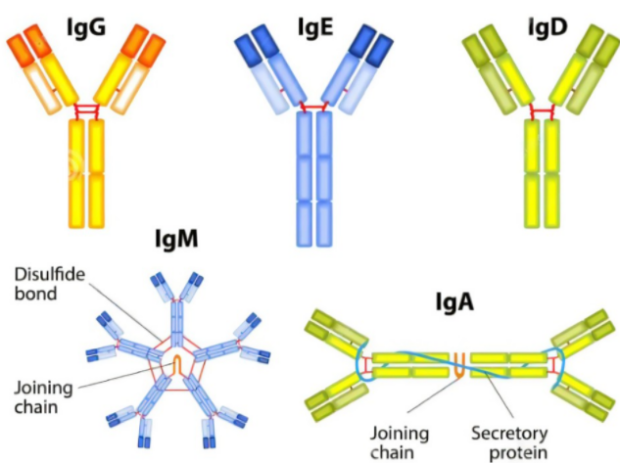
图2 抗体类别示意图
体细胞超突变(SHM)是另一种B细胞特异性反应,发生在CSR之后。一般体细胞基因突变率极低(1/1010-1/107),而在非常有限的B细胞染色体BCR V区基因片段中却能产生相当高频率的突变。在生发中心的活化B细胞每分裂一次,BCR的V区基因就会产生大约1/103碱基对突变。当遇到抗原时,抗原激活的成熟B细胞通过激活诱导胞嘧啶脱氨酶(AID)的活性启动SHM。
CH基因组转录过程中产生的转录泡、负超螺旋结构以及R-环产生的单链DNA招募激活诱导胞啶脱氨酶(AID)。AID将单链DNA S区上的胞苷(C)残基脱氨产生尿苷(U)。脱氨基的DNA被碱基切除、错配修复处理,发生双链DNA断裂(DSB)。双链DNA断裂使细胞内双链DNA损伤应答系统激活,最终非同源DNA末端连接系统将两个S区之间的 DNA 作为染色体外环丢失,完成DSB连接。
B细胞在外周淋巴器官生发中心接受抗原刺激后快速增殖,具有抗原亲和力低的B细胞通过在Ig可变区外显子部分引入点突变,对Ig基因进行修饰,以产生更高亲和力的抗体。
体细胞超突变可能会增加B细胞受体(BCR)对特定抗原的亲和力,当然也可能会导致BCR对抗原识别的丧失或产生自我反应性受体。大多数BCR亲和力降低甚至不表达BCR的突变B细胞克隆不能结合FDC(滤泡树突状细胞)表面的抗原,因无法将抗原呈递辅助性T淋巴细胞获取第二信号而凋亡;少部分突变B细胞克隆的BCR亲和力提高,表达抗凋亡蛋白而继续存活,从而实现了抗体亲和力的筛选。






南京德泰生物工程有限公司 Nanjing Detai Bioengineering Co.,Ltd. ©2024 All Rights Reserved
 苏公网安备32011202001300
苏公网安备32011202001300