
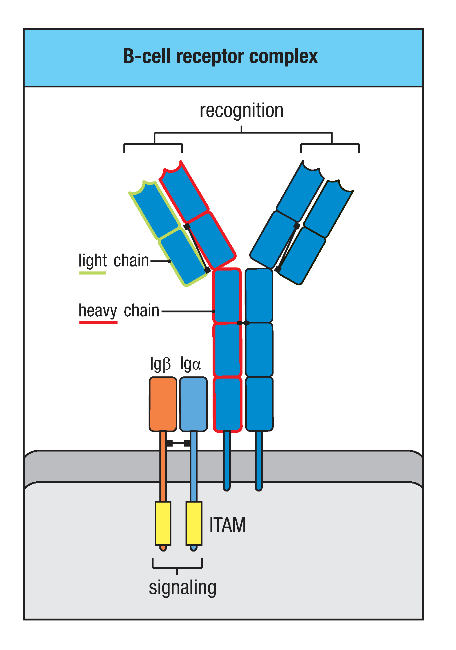
图1 BCR复合物结构
B细胞抗原受体(B-cell receptor, BCR)是一种位于B细胞表面的负责特异性识别及结合抗原的分子,其本质是一种膜型免疫球蛋白(mIg)。BCR具有抗原结合特异性,每个个体的BCR多样性高达5×1013,构成容量巨大的BCR库,赋予个体识别各种抗原,产生特异性抗体的巨大潜能。
BCR是由两条相同的分子量较小的轻链(L链)和两条相同的分子量较大的重链(H链)组成的四聚体结构。同分泌形式的Ig不同,BCR的重链是穿膜的多肽链。由于BCR的胞质区很短,不能直接将抗原刺激信号传递到B细胞内,因此完成BCR信号转导功能还需要Igα(CD79a)和Igβ(CD79b)的协助。Igα和Igβ都是免疫球蛋白超家族结构相关基因的表达产物,其在物种之间高度保守。Igα和Igβ都属于I型跨膜糖蛋白,在B细胞中以二硫键相连形成异源二聚体,并同mIg相连接。Igα、Igβ和mIg的跨膜区均有极性氨基酸,能够借助静电吸引而组成稳定的BCR复合物。Igα和Igβ的胞质区含有免疫受体酪氨酸激活基序(ITAM),能够募集下游信号分子,从而转导抗原与BCR结合所产生的信号。
BCR识别胸腺依赖性抗原(TD抗原)对B细胞激活有两个相关联的作用:(1)BCR可特异性结合抗原,产生B细胞活化的第一信号;(2)B细胞通过内化BCR所结合的抗原,并对抗原进行加工,形成抗原肽——MHCII类分子复合物(pMHC II),提呈给抗原特异性辅助T淋巴细胞(Th细胞)识别并使其活化。Th细胞活化后通过表达CD40L与B细胞表面的CD40结合,提供B细胞活化的第二信号。
蛋白激酶对酶和其他蛋白质的磷酸化是细胞调节其生化活性的普遍机制,具有调控迅速、反应可逆等优势。其中一个重要效应为募集胞质蛋白参与信号通路。例如,许多涉及信号转导的激酶是位于细胞膜内表面的,不能有效激活胞质中的靶蛋白;当受体活化,磷酸化的转接蛋白可以形成靶蛋白结合位点,招募细胞质中游离靶蛋白在激酶附近富集,继而依次被磷酸活化。
在抗原受体的信号转导过程中,由酪氨酸激酶反应产生的磷酸酪氨酸可以形成以一个蛋白质结合位点,即SH2结构域。这种结合位点在多种胞内信号转导蛋白质中发现,包括Src家族激酶。
1. BCR-Igα、Igβ介导的胞内信号转导
BCR在与抗原特异性结合后即启动B细胞活化的第一信号——ITAM上的酪氨酸磷酸化。B细胞内负责磷酸化ITAM的激酶有Blk、Fyn、Lyn三种,它们属于Src家族酪氨酸磷酸激酶。Src家族激酶是脊椎动物和其他动物中控制细胞分裂与分化相关的信号转导通路的共同成员。Src家族激酶可用过N端结构域与未磷酸化的ITAM低亲和力的结合,当抗原结合导致受体聚集时,受体相关激酶发生磷酸化并互相激活,使得Igα、Igβ尾部ITAM依次磷酸化。ITAM单个酪氨酸磷酸化可以使得其与Src家族激酶的SH2结构域结合,进一步磷酸化ITAM其他残基。Src家族激酶酶活性是由激酶结构域和C端区域的自我磷酸化调节的,这些区域均有调节性的酪氨酸残基。激酶结构中的酪氨酸发生磷酸化使其具有活性,在C端的酪氨酸发生磷酸化则抑制其活性。CD45可解除此抑制效应,是受体介导信号通路所必需的一步。
磷酸化的ITAM招募酪氨酸激酶(Syk)。Syk是BCR信号通路的关键激酶,含有两个SH2结构域,只有与B细胞受体的双重磷酸化ITAM结合之后才有酶活。
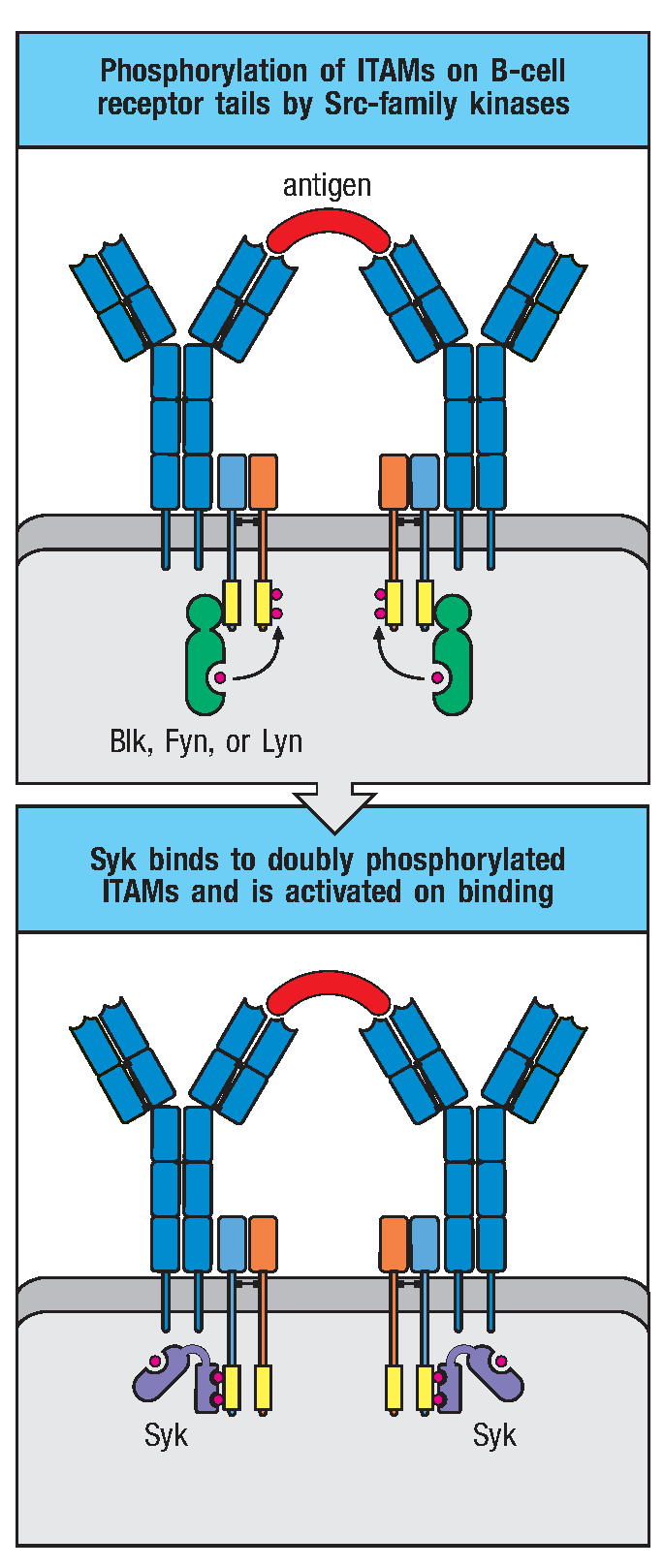
图2 BCR介导的胞内信号传导
Syk活化后使其他靶分子磷酸化,包括转接蛋白BLNK,由此招募Tec激酶,后者进一步激活磷脂酶Cγ2(PLC-γ2),催化细胞质内的磷脂酰肌醇二磷酸(PIP2)分解为三磷酸肌醇(IP3)和甘油二脂(DAG)。这两个分子都是信号通路中的重要组分。
IP3与内质网上的相应受体相互作用可引起Ca2+进入胞浆,内质网内腔的Ca2+浓度降低导致内质网膜蛋白STIM1与质膜上的钙离子通道ORAI1结合,促使ORAI1打开并让细胞外液的Ca2+流入细胞质,使信号得以维持。当细胞质中钙离子浓度增加时,会激活钙调蛋白(Calmodulin,CaM)使其构象改变。钙调蛋白可结合钙调神经磷酸酶(Calcineurin,CaN),CaN可以将NFAT(一种由钙离子通路调控的蛋白质,称为活化T细胞转录因子)上的磷酸化修饰水解,从而使其进入细胞核内启动下游基因转录。
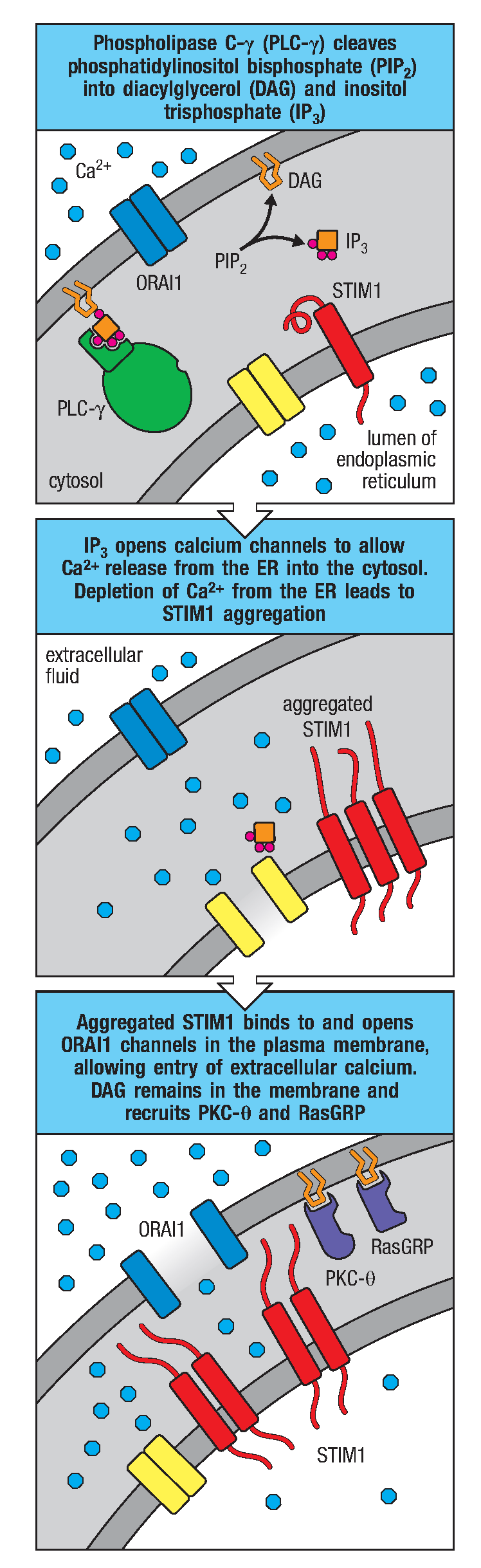
图3 钙离子信号通路
PIP2裂解的另一个产物为DAG,它可以保持与质膜内表面相联的状态,有助于活化蛋白激酶C(Protein kinaseC,PKC)家族成员。PKC家族大多为Ser/Thr蛋白激酶,可帮助启动通往细胞核的信号转导通路。另外,DAG也可招募RasGRP蛋白,RasGRP是Ras的一个鸟苷交换因子,含有C1结构域,用来激活附着于质膜上的Ras蛋白。Ras被激活后,下游再依次激活Raf、Mek、Erk三个激酶。因为Erk属于MAPK家族蛋白(mitogen-activated protein kinase),所以Ras激活产生的信号可传导到MAPK,MAPK再进入细胞核,磷酸化对应的转录因子。
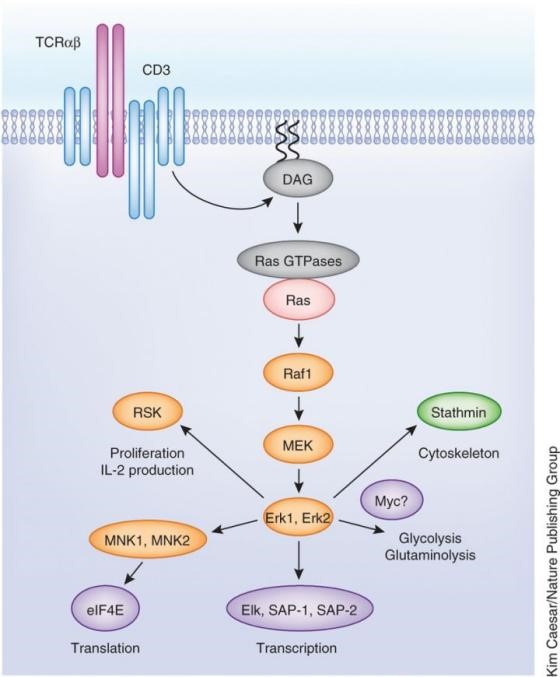
图4 DAG信号通路
最后通过蛋白激酶C(PKC)、丝裂原活化蛋白激酶(MAPK)及钙调蛋白等信号转导通路继续转导并最终激活NF-κB、AP-1、NFAT等转录因子,从而启动与B细胞活化、增殖、分化相关基因的表达。
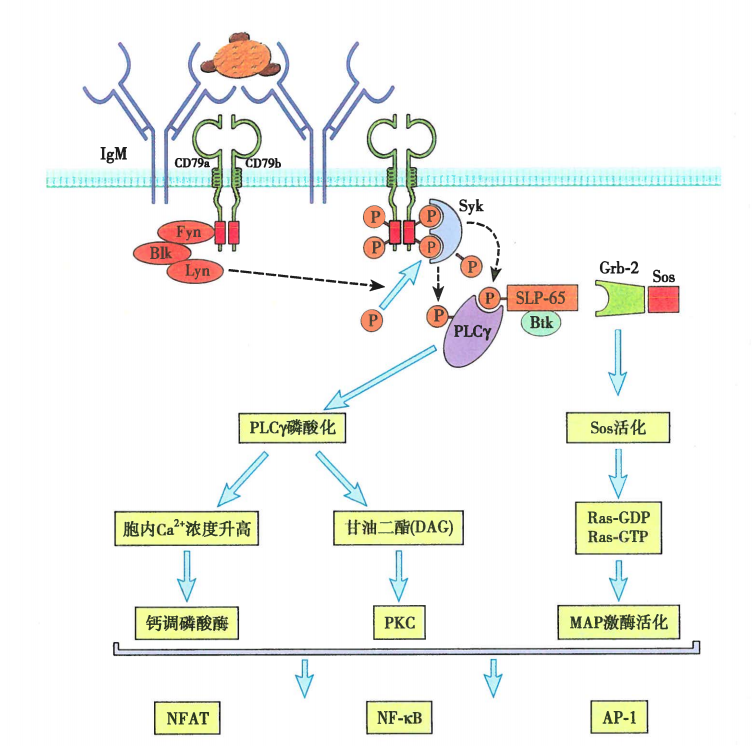
图5 BCR复合物介导的胞内信号转导
2. BCR共受体加强活化信号
B细胞表面的CD19/CD21/CD81复合物被称为B细胞共受体(B cell co-receptor),由CD19、CD21和CD81通过非共价键交联而成。复合物中,CD21可以和抗原结合从而增强BCR与抗原结合的稳定性,同时能识别与BCR-抗原结合的C3d,并通过交联CD19向胞内传递信号;CD19的胞浆区有多个保守的酪氨酸残基可以招募含SH2结构域的信号分子,与Igα/Igβ共同传递B细胞活化信号;CD81的主要作用可能是连接CD19和CD21,稳定B细胞共受体复合物。B细胞共受体通过稳定BCR与抗原的结合以及加强信号的传导,能够显著提高B细胞对抗原刺激的敏感性。
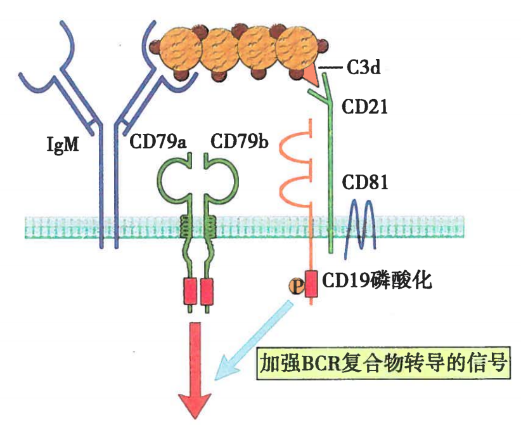
图6 B细胞共受体在B细胞活化中的作用
B细胞的第二活化信号又称共刺激信号,由Th细胞与B细胞表面多对共刺激分子相互作用产生,其中最重要的是CD40/CD40L。CD40表达在B细胞表面,CD40L表达在活化的Th细胞表面。CD40L与CD40相互作用,向B细胞传递活化的第二信号。
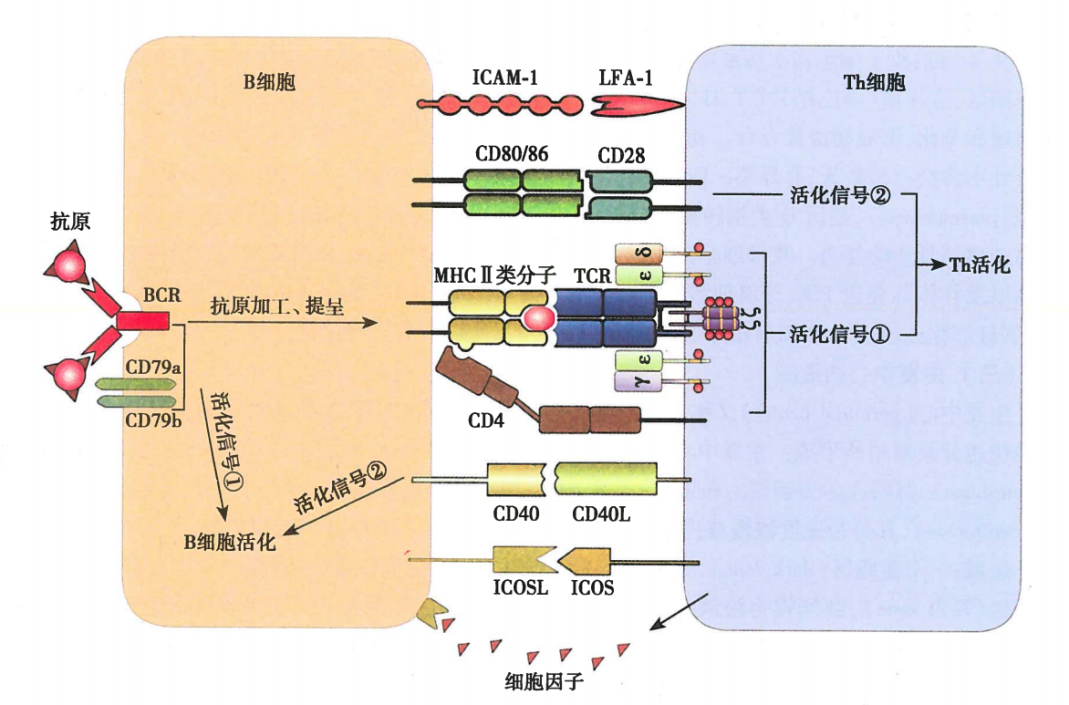
图7 B细胞与Th细胞相互作用






南京德泰生物工程有限公司 Nanjing Detai Bioengineering Co.,Ltd. ©2025 All Rights Reserved
 苏公网安备32011202001300
苏公网安备32011202001300