
双特异性抗体按结构区分主要有两大类:含FC区的双特异性抗体(IgG-like双特异性抗体)与不含Fc区的双特异性抗体(non-IgG-like双特异性抗体)。
IgG-like双特异性抗体结构与普通的单克隆抗体类似,呈”Y”型结构。具有较好的稳定性,较高的亲和力,在体内的半衰期比较长。
Triomabs双特异性抗体(Trifunctional Antibodies) 是将 CD3 特异性大鼠源 IgG2b 抗体和肿瘤靶向小鼠源 IgG2a 抗体进行体细胞杂交获得的双特异性抗体。Triomabs 通过 Fv 功能区分别结合肿瘤细胞及 T 细胞,通过 Fc 功能区募集表达 FcR的功能细胞,如 NK 细胞、单核细胞、巨噬细胞、粒细胞及树突状细胞等,形成复合体,刺激 T 细胞分泌细胞因子清除肿瘤细胞 因此 Triomabs 又被称为三功能抗体。

图1:Triomabs双特异性抗体结构及作用原理
杵臼结构抗体是利用 knob-in-hole技术制备的双特异性抗体。其结构特点为:组成双特异性抗体的两对轻重链对,其中一对的重链的CH3区发生突变形成一个突起的“杵”的结构,另一对的重链的CH3区发生突变形成一个凹陷的“臼”的结构,杵臼结构设计有利于两种种异源抗体重链的正确装配。

图2: 单克隆抗体、三功能抗体、knob-in-hole抗体结构
CrossMAb双特异性抗体是利用CrossMAb技术,将双特异性抗体的其中一条Fab抗体臂的功能区进行互换得到的双特异性抗体。

图3:常见的CrossMab双特异性抗体结构
CrossMAB是一种抗体Fab抗体臂的功能区互换的技术,是由罗氏开发的技术平台。该技术在“knob-in-hole”技术的基础上通过Fab臂功能区互换,解决了同源轻重链正确装配的问题,进一步提高了装配的成功率。
在正常抗体轻、重链的N 末端分别再接入另一个抗体的VL 和VH,形成双特异性抗体。
又称作DAF 抗体(Dual Action Fab),是普通抗体经过噬菌体展示技术改造得到的抗体,其结构特点为每一个抗原结合臂都有两种抗原的结合靶点。

图4:DVD-Ig与2 in 1-IgG结构图
这种构型的双特异性抗体缺失了Fc区,由两个抗体的VH区及VL区组成或者由Fab片段组成。此类双特异性抗体主要有 BiTE,DART,TandAbs,bi-Nanobody 等。
BiTE 是将抗 CD3 单链抗体( scFv) 与不同抗肿瘤细胞表面抗原的单链抗体(scFv)通过肽段进行连接而获得,可同时结合 CD3 阳性 T 细胞及肿瘤细胞并诱导T细胞靶向杀伤肿瘤细胞。

图5:BITE结构与作用原理
BiTE的分子量为55-60KDa,属于小分子,渗透性好,可以到达大分子抗体难以到达的部位与抗原发生结合。但亲和力较低,在体内的半衰期较短。
DART 双特异性抗体是由两条多肽链结合形成的异源二聚体抗体,其结构是将一个抗体可变区的 VH 和 VL 序列分别与另一个抗体可变区的 VL 和 VH 序列连接形成。此外,在两条多肽链的 C 末端引入了半胱氨酸,通过半胱氨酸形成链间二硫键,提高产品的稳定性。

图6:DART双特异性抗体结构
DART的作用主要有三方面:与靶向配体结合,实现细胞因子阻断;与靶细胞结合,实现抑制或阻断细胞激活信号;招募效应细胞杀伤靶细胞。

图 7:DART作用机制
双特异性抗体是四价的抗体分子,结构为 Fv1-Fv2-Fv2-Fv1,是由两分子肽链反向配对形成的同源二聚体分子。TandAbs 相对分子质量约为 110 kD,介聚体分子于全分子抗体及 BiTE之间。TandAbs可以同两种抗原结合,并且每种抗原都有两个结合位点。
TandAbs 可以招募效应细胞(T细胞或NK细胞),并对靶细胞(肿瘤细胞或癌细胞)起到杀伤效果。当TandAbs连接了T细胞与肿瘤细胞后,T细胞会被激活,释放穿孔素、颗粒酶、溶酶体酶等物质,这些物质会被传送到T细胞的细胞膜并分泌到细胞外的基质中。穿孔素会使靶细胞内形成气孔,从而促进裂解物质的进入,进而引起靶细胞的裂解。
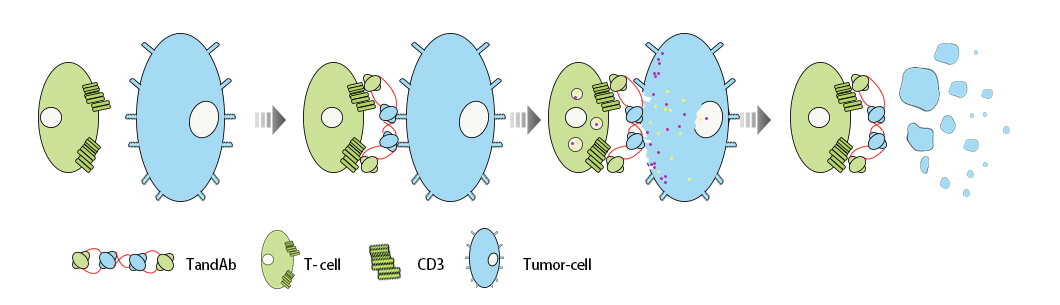
图8:TandAbs结构与作用原理
含Fc区的双特异性抗体:溶解性和稳定性好; 具有较长的半衰期;ADCC和CDC效应,增强肿瘤杀伤效果。
不含Fc区的双特异性抗体:体型较小; 改良后大幅降低使用的剂量,约为普通抗体的1/100以下。
引用:郭婷婷,梁锦峰.双特异性抗体药物的研究进展[J].中国新药杂志,2016,25(5):521-522.





南京德泰生物工程有限公司 Nanjing Detai Bioengineering Co.,Ltd. ©2025 All Rights Reserved
 苏公网安备32011202001300
苏公网安备32011202001300