
四个已知结构中的H1区长度相同,横跨于V区的顶部:
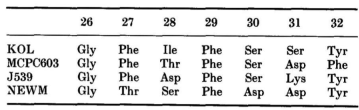
NEWM和MCPC603中H1的折叠非常相似,它们与KOL和J539中的折叠也很相似(图1)。
图1 VH结构域H1区构象
在观察到的H1结构中,位于26位的甘氨酸产生了一个角度较大的转角,其ϕ、φ值(+75, 0)超出了非甘氨酸残基所允许的范围。位于29位的苯丙氨酸深埋在框架区中,与残基34的侧链以及残基72和77的主链相邻。27位的残基(Phe或Thr)部分埋在残基94旁边的表面空腔中。在这四种结构中,位于26、34和94位的残基相同或相似,分别是Gly、Phe、Met/Tyr和Arg。
根据Kabat等列出的185个H1区已知的人类和小鼠VH区序列。在178个序列中,170个与已知结构中的序列长度相同,其余1个小鼠序列长一个残基,6个人类序列长两个残基。
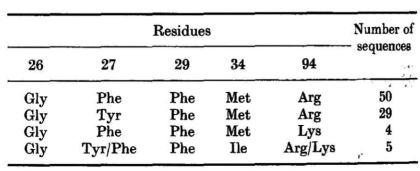
在170个H1为7个残基的序列中,有115个94位残基是已知的。其中,四分之三在26、27、29、34和94位的残基与已知结构中的残基相同或非常接近:
环状结构的长度和H1与框架包装相关位点残基的保守意味着,至少在这些VH结构域中,H1的构象与已知结构中的构象接近。
H2区连接位于β-片层相邻链的框架残基52和残基56。在已知的VH结构中,H2环的长度各不相同:在NEWM中,它包含三个残基;在KOL和J539中,它包含四个残基;在MCPC603中,它包含六个残基。Kabat等列出了127个已知整个H2区序列的人类和小鼠VH序列。除一个序列外,所有序列中H2的长度都与已知结构之一相同:

42个H2区6残基的序列都属于小鼠的亚群Ⅲ。
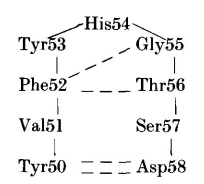
NEWM H2区域的三个残基(Tyr53、His54和Gly55)构成了一个七残基转角的顶点。转角中的其他四个残基来源于框架区的一部分:
NEWM中发现的构象几乎和上文所述七残基转角的构象一致,在第五个位置(即NEWM中的第55位)有一个Gly,Asn或Asp。根据Kabat等列出的13个三残基H2区域中,有9个在55位有Gly残基,4个有Asp,预测这些H2区具有NEWM类似的构象。
J539和KOL中的H2区域(52a至55)形成四残基转角:

这些转角的构象由甘氨酸残基的位置决定。J539中的H2具有最常见的四残基转角构象:前三个残基处于近似αR构象,第四个残基(Gly55)处于αL构象。KOL中的H2则不同:Gly54处于αL构象,其他三个残基处于αR构象。
71个H2区为4个残基的序列,有10个Gly、Asn或Asp残基出现在54号位置,12个出现在55号位置,32个同时出现在54号和55号位置。在那些仅在54位有Gly、Asn或Asp残基的序列中,预测会出现类似于KOL中的H2构象。在55位有Gly的情况下,预测类似于J539中的H2构象。
MCPC603 H2区域的六个残基是十残基发夹转角的一部分。目前,实验和理论方面的证据不足,无法制定出这种大型转折的规则构象。因此不知道小鼠亚群Ⅲ中的其他六位残基H2区域是否与MCPC603中的构象接近。
H3区域由残基96至101组成。VH结构由三个基因重组而成:VH(编码残基1至94或95)、D(编码1至13间残基)和JH。
人类的JH种系基因有6个,分别由以下氨基酸序列编码:

小鼠的JH种系基因有4个,由以下氨基酸序列编码:

由于D和JH基因的连接末端可能会发生变化,因此在JH基因开头编码的残基可能不会出现在最终结构中。体细胞突变会导致该区域序列的进一步多样性。因此,J539、NEWM、MCPC603和KOL的H3区域在长度(分别为6、7、9和15个残基)、序列和构象上存在很大差异也就不足为奇了。在此,我们将只讨论MCPC603的H3区,因为分析表明,其构象至少部分存在于其他几种免疫球蛋白中。
MCPC603中的H3区域形成了一个大的发夹环:
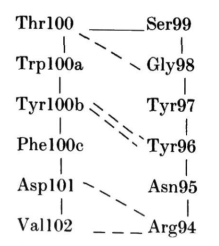
对于这种大环,ф、φ值范围将允许多种构象,而实际发现的构象将取决于与蛋白其余部分的堆积情况。在MCPC603中,H3的构象主要由VH结构域内和VL-VH界面的Arg94、Tyr100b、Phe100c和Asp101残基的相互作用决定。残基96至100a的侧链位于蛋白表面。
Arg94穿过H3发夹,与VL-VH界面的Asp101、Tyr100b和Phe100c形成表面盐桥(图2)。相当于100c位置上的残基通常是VL-VH界面保守核心的一部分,在83%的已知序列中,该位置的残基为Phe或Leu。在MCPC603中,Tyr100b与VL的Tyr49相邻,形成一个大空腔,两个Tyr残基的羟基都位于接触表面。在82%的VL结构域中,49为Tyr或Phe。相当于100b位置上的不同残基可产生不同的H3构象。例如,在KOL中是Gly,而与VL Tyr49相邻的空腔则由相当于100a位置的Phe填满。这使得KOL中H3的构象与MCPC603中的构象截然不同。
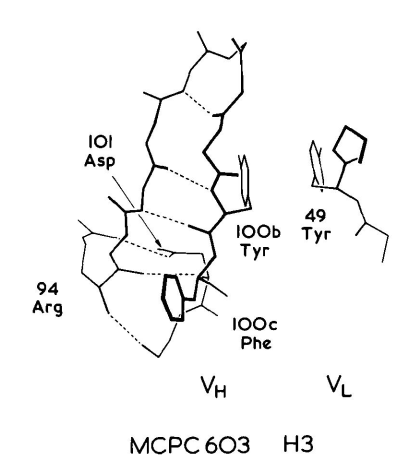
图2 MCPC603 H3区构象
位于100-100-101位的Tyr-Phe-Asp序列存在于人类基因JH2和JH4以及小鼠基因JH1和JH2中。人类JH5基因用Trp取代了Tyr。如果这些残基没有在基因重组或体细胞突变过程中丢失,通常认为这些J4基因会产生与MCPC603中的H3构象接近的H3构象。根据Kabat等序列表中查找出的长度至少为6个残基的H3区域,这些区域94位为Arg残基,101位为Asp残基,101位之前的两个位置有Tyr-Phe,即具有以下形式的区域:
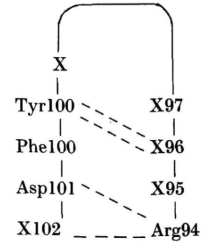
根据Kabat等列出了的28个人类和77个小鼠序列的整个H3区域。其中,1个人类序列和48个小鼠序列完全符合长度和序列条件。另外5个人类序列很接近,它们的区别仅在于94位为Lys,Phe或Trp取代了Tyr,或Met取代了Phe。这54个序列中H3区域的长度分布:

在这些序列中,预测H3具有与MCPC603中相同的构象。
上文对超变区的描述表明,它们的主链构象完全由每个区域内的特定残基决定。实际上,这些构象可能会受到环境的影响。对特定区域的影响可分为两部分:构象的局部变化和结合位点中相对位置的变化。
衡量两种肽构象差异的标准是它们最佳叠加后原子位置的r.m.s.差异。在上面的章节中,我们报告了具有相同折叠的不同免疫球蛋白结构的超变区主链区域的r.m.s.差异。r.m.s.差异很小:在高分辨率下测定结构的r.m.s.差异小于0.5 Å,在中等分辨率下测定结构的r.m.s差异通常为1.0 Å或更小,这主要是由于肽段取向不同造成的。只有在H1区才会有显著差异,尽管构象上的差异仍然很小。
为了确定免疫球蛋白结构中超变环相对位置的差异,我们进行了以下计算:将Fab蛋白NEWM、MCPC603、KOL和J539 VL-VH框架残基拟合,将REI和RHE VL框架残基拟合。框架叠加后,我们计算了叠加同一折叠的超变区所需的额外位移,例如J539、REI和MCPC603的L1区的共同残基。
计算结果见图3。在Fab蛋白中,同一折叠的超变区在位置上相差0.2 Å至1.5 Å。出现这些差异的部分原因是,尽管VL-VH二聚体具有相同的残基接触模式和非常相似的堆积几何形状,但VH相对于VL的取向存在微小差异。
REI VL结构是通过Bence-Jonex蛋白确定的。它包含一个VL-VL二聚体,它们在REI中的排列与Fabs中的VL-VH排列非常相似。REI超变区相对于框架区的位置与Fabs中的位置相同。

图3 同源超变区相对于β-片层框架的位置差异
实线表示Fab结构中超变区之间的差异,断线表示Bence-Jonex蛋白和Fab超变区之间存在的差异。
因此,在这些结构中,超变区环境的不同只会导致主链构象的微小差异,其相对于框架的位置差异也不会超过1.5 Å。Bence-Jonex蛋白RHE和MCG中,超变异区的环境与免疫球蛋白中通常存在的环境截然不同。RHE中VL-VL二聚体的堆积方式与VL-VH二聚体的堆积方式截然不同,因此其超变异区的环境也与本文讨论的其他免疫球蛋白的环境截然不同。这些环境差异对RHE中L1、L2和L3的构象影响不大:它们与Fab KOL中的同源区的坐标r.m.s差小于0.3 Å。相对于框架区,它们确实会对L1、L2和L3位置产生一些影响:RHE中的位置与Fab中的位置相差可达2.2 Å。
在Bence-Jonex蛋白MCG的结构中观察到更复杂的情况,这种蛋白的晶体具有不对称单元组成的二聚体,两个VL单体处于不同的环境中。其中一个单体的L1区处于螺旋构象,与预期一致;而另一个单体的L1区则由于残基3至32与邻近分子的紧密接触而无法具有螺旋构象。密切接触产生无序结构表明,L1区域只有有限的灵活性。
上文中,我们将可变区的两个部分进行了精确的结构区分:保守的β-片层框架和主链可变区构象。这两部分对抗原结合位点有何贡献?
超变区聚集在VL-VH二聚体的一端,形成一个表面,其中一部分与抗原相互作用。图4显示了MOPC603中这一区域的空间填充图。在蛋白的这一部分,溶剂可接触到的VL残基为27至32、49至53和92至94,在VH中为28至33、52至56和96至100a。第3节中定义的β-片层以外区域在VL中为26至32、50至52和91至96,在VH中为26至32,52至56和96至101,溶剂可接触区域与其最多相差两个残基。在表6中,我们列出了J539、KOL和NEWM形成相同区域的溶剂可及残基。KOL、NEWM、J539和MCPC603中溶剂可及残基非常相似,但并不完全相同。环长度和序列的变化可能会导致环末端的一两个残基被掩盖或一两个框架残基暴露。
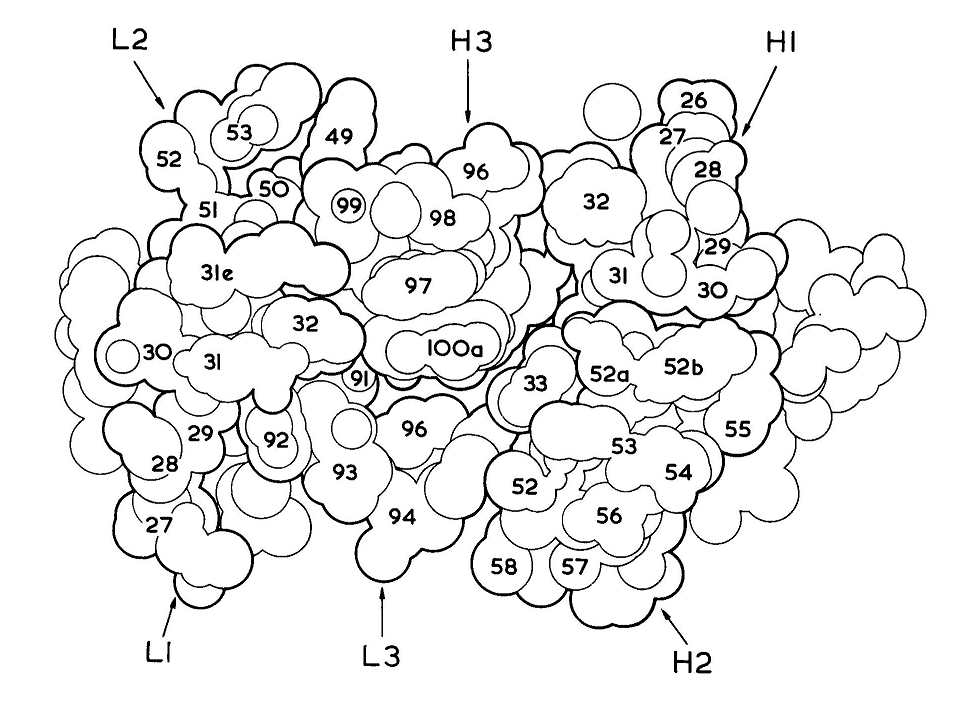
图4 MOPC603高变区空间填充图
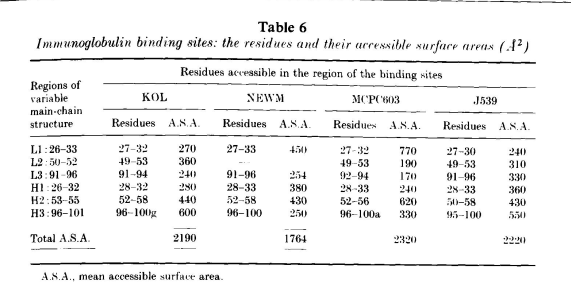
除H2外,表6中列出的区域与根据序列变异性确定的互补性决定区域(CDR)相似。表6中的H2包含残基50至58,相应的CDR包含残基50至65。Padlan发现,此CDR的前三个残基和后六个残基与NEWM和MCPC603具有相同的结构。残基59至65位于VH结构域的一侧,与其他超变区的距离相当远。残基59至65的侧链可以被溶剂接触到,其序列的变化可能仅仅反映了结构和功能约束的缺乏。
表6还列出了构成结合位点的各环状结构的可接触表面积。KOL中的H3不同寻常地大,有15个残基,对总表面积的贡献最大。而J539、NEWM或MCPC603中的情况并非如此,中等大小的H3区域与其他环的贡献相似。H3在抗体特异性中的重要作用不是来自于它的大小,而是来自于它在结合位点中的中心位置。
在J539、KOL和MCPC603中,超变异环总溶剂可接触表面积约为2250 Å2:而在NEWM中,由于未包含L2,其可接触表面积为1760 Å2。对寡聚蛋白的分析表明,在分离状态和结合状态的结构非常相似的情况下,稳定的结合是由表面积小于这些结合位点总表面积的表面形成的。通常情况下,单体有500 Å到1000 Å埋藏在界面中,占超变环可接触总表面积的四分之一到一半。这些界面中氢键和盐桥的数量各不相同。
对免疫球蛋白D1.3和鸡蛋蛋清溶菌酶形成的复合物的研究支持了抗体与蛋白之间的作用会涉及到与寡聚蛋白中的相似表面这一猜想。这种结合不涉及主链构象的重大变化。与溶菌酶接触的抗体残基在VL中为30、32、49至50、91至93,在VH中为30、32、52至54和96至99。互作界面由690 Å2的抗体表面和750 Å2的酶表面组成。
在这篇论文中我们试图识别决定高变区构象的残基。我们提出,如果我们定义的残基存在于其他免疫球蛋白序列中,那么它们的高变区将与已知结构的高变区具有相同的构象。对免疫球蛋白序列的分析表明,大多数高变区具有一组或多组主链构象。我们称这些构象为“典型结构”。
Vκ结构域AU和ROY的原子结构支持了我们分析的一些结论,这两种蛋白的序列与REI的序列分别有18和16个残基不同。其中8个位置的变化发生在高变区。

在30,31,93和96位的残基变化涉及体积和化学性质的巨大差异。然而,从上面给出的分析,我们认为它们应该产生与REI中相似的主链构象,事实上也是如此。
为了测试分析的准确性,我们用上述结论来预测新的免疫球蛋白的可变结构域。预测在通过X射线分析确定结构之前进行的。
基于本文所述工作的预测方法与前人所使用的方法有根本区别,之前的做法是将其免疫球蛋白高变区序列与已知结构中相应高变区序列进行了比较,然后从长度和总体序列同源性最接近的区域建立了每个环的模型。在某些情况下,调整是为了适应序列的差异。
基于本文所述工作的预测方法,如为了确定L1的构象,我们将检查第2、25、29、30、33和71位的残基。如果在这些位置上发现的残基与上文列出的一组残基相匹配,那么无论其他位置上的残基是什么,我们都会认为L1具有相应的典型结构。如果这些位置上的残基与其中一组残基不匹配,那么无论超变区其余部分的同源性多么接近,我们也无法鉴定为已知的典型结构之一。
我们对免疫球蛋白D1.3的结构进行了预测,并在确定其结构之前将预测结果发送给了进行X射线分析的小组。主链的构象是通过本文描述的分析方法预测的;对于侧链的构象,我们使用了之前描述的程序。在预测之后,我们根据2.8 Å电子密度图确定了D1.3的原子结构,并对框架结构和六个超变区中进行了预测。结果证实了晶体结构分析与预测已知结构相同。
D1.3的三个超变区(L1、L2和H2)与已知的典型结构长度相同,晶体学分析证实了这三个区域的折叠接近典型结构的预测。其他三个超变区的序列与已知的典型结构相同或长度相似,但在决定构象的位点上,它们的残基相似但不完全相同。在预测这些区域的结构时,我们必须判断这些差异是否会产生不同的主链构象。在其中的H3中,我们的预测是正确的,而在另外两个L3和H1中,我们的预测部分是错误的。
综上,我们确定了在已知结构中造成超变区构象的残基,如果这些特定残基出现在其他免疫球蛋白中,它们的超变区也将具有相同的结构。这也表明,当残基不完全相同时,即使变化很小,也很难有把握地预测结构。
我们对免疫球蛋白序列的分析表明,它们中许多超变区形成了已知结构的六个VL结构域或四个VH结构域中的典型结构之一。大多数超变区具有上述主链构象之一,这一结论对H3的适用可能有限,因为H3在长度和序列上的变化远大于其他区域。然而,我们的分析确实表明,在已知的小鼠序列中,有一半的H3区域的构象至少部分接近于MCPC603中的构象;D1.3中H3的成功预测证实了这一点。
对其他抗体晶体结构的分析将扩大典型结构的范围。尝试预测更多的结构,并在晶体学结构确定后对其进行测试,将提高我们理解负责其构象的残基可能发生的变化所产生的影响的能力。对抗体结构的预测不仅有助于测试我们对负责典型结构构象的残基的识别是否准确。它对于设计具有特定特异性的抗体也至关重要。
参考文献
Chothia,C. and Lesk,A.M. (1987) Canonical structures for the hypervariable regions of immunoglobulins. J. Mol. Biol., 196, 901–917.






南京德泰生物工程有限公司 Nanjing Detai Bioengineering Co.,Ltd. ©2026 All Rights Reserved
 苏公网安备32011202001300
苏公网安备32011202001300